4. 遺伝子研究の技法について
(1) PCR法:
DNA断片を痕跡から大量に増幅する方法である PCR法(ポリメラーゼ連鎖反応法・polymerase chain reaction、1985~)が発見されたため、DNAの調査(配列決定、親子鑑定・犯罪捜査、・・)、遺伝子組み換えなどの遺伝子工学が飛躍的に発展した。(それ以前は、たとえば ヒトのミトコンドリアDNAを採取するために、産婦人科の協力を得て ”後産”をミキサーですり潰して採取していた。)
PCR法は、鋳型DNAから、ポリメラーゼという酵素を用いて 必要とする短い断片(プライマーで指定したところからの部分)のみを複製し、これを繰り返して大量のDNA断片を作製する方法である。 また現在は、long PCR として 10kbp以上の複製が可能となっている。 鋳型DNAの抽出は、細心の注意が必要で、違うもの(コンタミなど)を含まないように注意しなければならない。
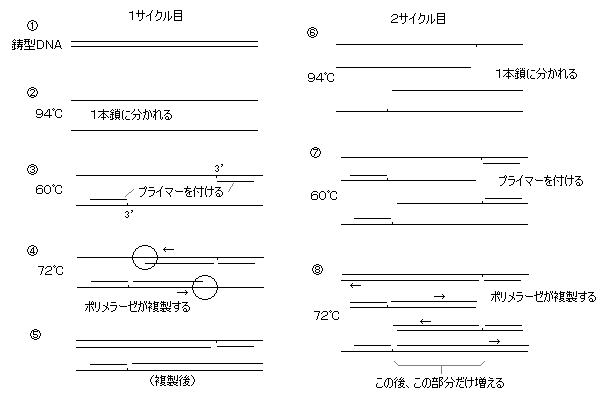
(以下、手順)
まず、複製するDNAの それぞれの鎖の3’(開始点)の外側20個程度の塩基配列をデータベースなどから決定し、それぞれの相補的な塩基配列のプライマーを化学的に合成する。(業者が合成してくれる。通常20塩基、最大50塩基程度。普通は脱塩・未精製品で充分)
自動温度制御槽(PCリアクター)に、鋳型DNA(テンプレート)、プライマー、耐熱ポリメラーゼ(好熱菌の耐熱性DNAポリメラーゼ(Taq ポリメラーゼ など)使用、高温でも分解しないので一度加えればよい)、dNTPs(4種の、ヌクレオチド(A、C、G、T)・3リン酸 の混合物)、バッファー溶液(MgCl2含む)を加えスタート。
高温(94℃、30~60秒)にすると、鋳型DNA2本鎖が熱で変性して、1本鎖に分かれる。
60℃(プライマーによって若干前後する)に急冷し、プライマーを1本鎖になったDNA上の必要とする部分の両端に付ける。(プライマーのアニーリング)
再び温度を 60~72℃の設定温度(ポリメラーゼが働きやすい温度)に上げ、1~2分保つと、プライマーで挟まれた部分の2本鎖DNAができる。
これで1サイクルであって、特定の部分が抽出されるまで最低3サイクル必要であり、最初の2サイクルではそれぞれ片端の長いDNAができるが、20~30サイクル程度繰り返すので、最終的には 必要な部分の短いDNAが大部分となる。(数十万倍に増幅)
増幅されたDNAは、ゲル(ポリアクリルアミドゲル)に入れ 電気泳動で分離、あるいは、キャピラリ電気泳動法(毛細管により対流を防ぐ、現在こちらの方が主流)で分離し、UV、発色剤などで確認する。
* 塩基配列の決定法は、シークェンシング(それぞれ4種の塩基の所で切断し、それぞれの長さの断片を 短い方から並べなおし、それを切断した塩基で読むと、もとの配列になる)、酵素法(PCR法で、特定の場所にプライマーを付け、そこを起点としてポリメラーゼで複製するときに、4種のうちの特定の塩基のところで複製が止まるように反応系を定める)、化学分解法などがある。
(2) 人工ゲノムの合成技術(?)
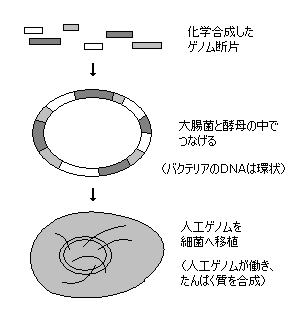
DNA生命体は、エントロピー増大側により、”自然発生的”には到底 作られるわけが無いが、人工的には、すでに存在する生物体や生物素材をふんだんに用いての話であるが、先の”ドリー”の例もある通り、ある程度 加工は可能となってきている。
・・・ 神様による偉大な創造のわざに比べると、それは”食品加工”(組織の分解、加熱調理など)に”毛が生えた”レベルである。人間ができることは、どこまでいっても、マクロの操作(混ぜる、熱処理する、電気をかける)と、それによってできた混合物をなんとか分離する、といった程度である。
また、不確定性原理によって、ミクロのものを直接扱うにも限界がある。顕微鏡で見るだけで、どこかにすっ飛んでしまう。
今年(2010年)5/20 Science誌に、塩基配列をデザインした”人工ゲノム”を移植して異種細菌が”変身”し、DNAを設計した元の細菌のたんぱく質を別細菌が合成した、というニュースが公表された。(J・クレイグ・ベンター研 Daniel Gibsonら・米メリーランド州)
研究ではまず、牛の感染症(牛肺疫菌)を起こす細菌”マイコプラズマ・ミコイデス”の解析されたゲノムデータに基づき、この各部分の短いDNA断片を化学合成した。この化学合成のやり方は、(1)のPCR法で用いられる プライマーの合成法(50~塩基程度)として確立されている。分子量が小さい場合は人工的な化学合成が可能。
今回のポイントは、この短い断片群を 大腸菌、および、酵母に中に入れ、断片をつなげて長大な”人工ゲノム(?)”を完成させることである。
ただし、長いDNAのうちのすべての遺伝子ではなく、そのたんぱく質を合成させる遺伝子部分がたまたま生き残り、移植細菌では他の遺伝子14個が消失または破壊されたそうである。
そして、その混合物をミコイデスと類似する別種の細胞に移植した結果、運良く生き残った”その遺伝子部分”がミコイデスのたんぱく質を合成した、というレベルのものである。
・・・・・ この技術は、「遺伝子組み換え技術」の一つであり、大腸菌、酵母の中の酵素群を利用するもので、純粋に”人工的”とは言えず、この DNA断片を設計どおり”つなげる”ということが、生物体を利用しなければできないところである。
また、DNAの全解析はできるが、そのデータの意味するところをすべて知っているわけではない。ましてや、データを「創造」することはできない。 すでに自然界に有るものを、機械的に”模倣”しただけである。
まだまだ、”人類史上初めて、「生命の創造」に手が届く段階に達した”、と言えるものではない。
この技術を応用すれば、望みのゲノムを設計して微生物に組み込み、現存しない”新種”を生み出せる可能性がある。将来、この手法を使ってこれまでにないゲノムを設計し、空気中の二酸化炭素から効率的に燃料の炭化水素を作り出す藻類や有毒物質を分解する細菌の合成、ワクチン生産の効率化といった応用 が想定されるという。石油大手エクソン・モービルはバイオ燃料を大量に生産する藻を作るため、この研究に資金を提供。製薬大手ノバルティスもワクチン開発のスピードアップに利用しようと研究を始めたそうである。しかし、強力な病原菌が開発され、テロに悪用される危険や、自然界に無い生物が実験室から拡散する可能性があり、議論を呼んでいる。
(3) 発生の生化学:
動物の発生(胚発生)は、次のステップで行なわれる。
① 卵割(受精卵が細胞数を 2n で増やす)
② 胞胚期(ほうはいき、細胞数が増えると空洞ができる。ウニはこの時期孵化する)
③ 原腸の形成(胞胚が内側に陥入(かんにゅう)し、袋状(刺胞動物、扁形動物の場合)または管状の消化管となり、一つの口を作る。この口は、軟体動物、節足動物、環形動物、多くの無脊椎動物では口、 棘皮動物、脊椎動物では肛門となり、それぞれ反対側に肛門と口ができる)

④ 胚葉の分化(内胚葉は消化管、 外胚葉は 表皮と神経、 中胚葉は 筋肉、血管系、体腔 が分化する)
⑤ 形態形成(ここから後は各器官が形成される段階になる。形態形成は 遺伝的な「プログラム」によって調節され、さらに環境要因により変わり得るものとなる。転写因子たんぱく質とDNAから生じた2次的遺伝子は、制御カスケード内のさらに別の遺伝子の発現を調整する。)
* 培養細胞 と がん細胞は分化は見られるが、形態形成はしない
誘導は分泌性因子を介した細胞間相互作用により行われると考えられるが、しばしば多数の因子が複雑な制御系を形成しており、分子メカニズムが解明されていないことも多い。(発生コントロール遺伝子、ホメオティック遺伝子、ホルモン、受精卵を縛る外力など)
* 発生の様子のスケッチは 古くから描かれてきたが、あたかも”胚発生の通りに進化した”かのように、”進化論的に”大いに誤解されて用いられてきた。特に、”個体発生は系統発生を繰り返す”という”反復説”を作り上げた ヘッケルは、自説を主張するために、故意に偽のスケッチを公表した。((→ 左図) 教科書にも載っているので、早急に修正しなければならない!) これは、実物のヒトの胎児のスケッチ(→ 右図)と異なる。
近年でも「個体発生は系統発生の反復はしないが、発生の初期ほど進化的に より古い形質が現れる傾向にある」といった考えは根強い。
(* ヘッケルの反復説の社会的影響は、ダーウィニズムと同様、曲解に近い形で社会的広がりを見せた。たとえば子供は大人にくらべて進化的に前の段階であると か、いわゆる原始的種族は、進化の段階が低い状態にあるといった拡張がおこなわれ、ナチス・ドイツなどの人種差別に利用されることもあった。)
胚発生の初期の様子が、一見、いわゆる”進化”のように見えるのは、「神様」が、その偉大な創造性を発揮して、より下等な動物を「試作」しながら 改良を加え、最終的にはヒトを造られたからである。 異種間の交配はできず、創造の初めから現在まで 種は独立している。ミッシングリンクは化石にさえも存在しない。
(「第5日: 海の巨獣・水の生き物、鳥 、第6日: 陸上動物、人間」(創世記 1:20-28)
さらに、近年の、生化学的、分子生物学的なアプローチによって、真の発生の過程が、スケッチなどの見た目の変化とかなり異なることが徐々に明らかになっている。
たとえば、目の発生過程(”進化”ではない!)について:
① 胚の原口背唇(一次形成体)が 外胚葉に働きかけ、神経管を誘導、脳に分化する。
② 脳の両側に眼胞(がんぽう)、次に、眼杯(がんぱい、二次形成体)となる。
③ 水晶体(三次形成体)、表皮に角膜を誘導する。

この目の働きを、解剖学的に説明するのはもはや不十分である。 (ヒトの網膜の光受容体のみについてであるが、)生化学的に説明すると、いかにその働きが複雑きわまるものであるかが分かる。これは、非常によく設計された「生化学的機械」の一つであり、この”設計図”は、すべてDNAの中にあらかじめ書き込まれているのである。(以下、説明文)
まず光が網膜に当たると、1個の光子が11-シス-レチナルと呼ばれる分子と相互作用し、数ピコ秒以内に トランス-レチナルに変化する。(1ピコ秒≒ヒトの髪一本の幅を光が駆け抜ける時間) レチナルの形が変化すると、固く結びついているロドプシンというたんぱく質の形を変化させ、メタロドプシンⅡとなって、トランスデューシンと呼ばれる別のたんぱく質と結合する。トランスデューシンは GDPと固く結合していたが、メタロドプシンⅡと相互作用すると、GDPが抜け落ち GTPが結合する。
この GTP-トランスデューシン-メタロドプシンⅡは、細胞の内膜に位置するたんぱく質の フォスフォジエステラーゼと結合する。フォスフォジエステラーゼは、メタロドプシンⅡが存在すると、cGMPという分子を切断する能力を持つようになる。細胞中に多くのcGMPが存在するが、この作用により、その濃度が下がる。 cGMPと結合するもう一つの膜たんぱく質は、イオンチャンネルであり、通常は細胞内のナトリウムイオンの数を調節する通路の役割をしている。通常はナトリウムイオンを細胞内に流入させているが、一方、別たんぱく質がナトリウムイオンを外へ汲み出していて、一定の濃度に保っている。 cGMP量が下がると、イオンチャネルは閉じ、正電荷を帯びたナトリウムイオンの細胞内濃度は下がり、細胞膜内外の電位差が不均衡になり、視覚神経を通して脳まで伝達される電流を引き起こす。
また、これを回復させる働きも存在する。暗闇では、ナトリウムイオンのほかにカルシウムイオンをも細胞内に流入させていて、カルシウムも別のたんぱく質によって汲み出され、一定の濃度に保たれている。cGMP濃度が下がると、cGMPを切断するフォスフォジエステラーゼの働きが遅くなる。 また、カルシウムイオン濃度が下がり始めると、グアニル酸シクラーゼがcGMPを再合成し始める。 さらに、ロドプシンキナーゼという酵素によって修飾を受けたメタロドプシンⅡは、アレスチンというたんぱく質と結合し、トランスデューシンをそれ以上活性化することができなくなる。 したがって、これらの作用により、1個の光子によって始まった信号の増幅機能を制限することになる。
トランス-レチナルは、最終的にはロドプシンを低下させ、11-シス-レチナルに戻り、次の視覚サイクルの開始点に戻す必要がある。それは、トランスレチノールが酵素によって水素原子が2個多く付いた形となり、その後次の酵素が11-シス-レチノールに戻し、3番目の酵素が脱水素して 11-シス-レチナルに戻すことによる。

このように、単純な水晶体や、単純な網膜、涙管、まぶたなどを作り出すことが、どれほど複雑で難しいことであるか、生化学的に見れば分かるのである。 同時に、これらは、一つでもその要素が欠けると全く機能しないものである。(DNA配列が突然変異して徐々にそうなった、という考え方はできない。)
したがって、それぞれの生体機械の各動作部分は、”これ以上単純化できない複雑さ”を持ったシステムであると言える。
(4) 発生 と 不完全性定理:
さて、20世紀に明らかにされた”人間が知る限界”の一つに、不完全性定理がある。 K.ゲーデルらによって証明されたこの定理は、グレゴリー・チャイティンによって、「ランダム性」についても成立することが証明された。
01010101010101010101 という数列は 01 を10回繰り返すという”圧縮可能”な数列であるが、たとえば、 10101110110011111010 という数列はこれ以上”圧縮不可能”であり、この数列は「ランダム」であると言う。 圧縮不可能とは、”これ以上単純化できない複雑さ”である。すると、
① ゲーデル・チャイティンの不完全性定理: 「任意のシステムSにおいて、そのランダム性を証明不可能なランダム数Gが Sに存在する」
・・・・ システム S の公理 と 推論規則 を記述するプログラムを 2進法にコードしたものが n ビットであるとすると、Sは、n ビットよりも長いランダム数(これ以上圧縮できない)を プログラムで表現できないので、Sは、n ビットよりも長い2進法数列でコードされる 自然数のランダム性を 証明できない。(計算できない)
また、ある公理系内で無作為に一つの命題を選び出したとき、その命題がその公理系内で決定(証明又は反証)可能である確率 を 「真理性 Ω」とおくと、
② チャイティンの定理: 「真理性 Ωは、純粋にランダムである」
・・・・ その命題を導出する方法が一切存在しない。命題が真であるか偽であるかは、コイン投げの結果と変わらない。”神様は数学においてもサイコロを振る”ことになる。(人が知らないだけ)
ここで、4つの塩基の暗号列であるDNAをコンピューターのプログラム(4進法・プログラムの総数=重複順列=4N個 → 2進法・2n個 に変換)とし、生物の発生過程をそのアルゴリズム(生化学的なすべての制御カスケード)とおく事を考える。
すると、複雑さがある程度以上になり、アルゴリズムがプログラムの長さ(=nビット)を超えると、生物の発生過程を”機械論的”に解釈するのに十分な発生過程を特定することができなくなる。
すなわち、システムにインプットした情報量以上の情報を、システムはアウトプットできないから、自己の情報量を超えたランダム性を決定できない。 したがって、アルゴリズムがそれ以上複雑に成長すると、原理的に、遺伝子過程の特定が不可能になる。
このことは、次のみことばに書かれている。
「夜は寝て、朝は起き、そうこうしているうちに、種は芽を出して育ちます。どのようにしてか、人は知りません。」(マルコの福音書 4:27)
・・・ みことばの種の成長と同じ。 良い麦と毒麦とは成長途中で見分けられない。人は、それが実を結んでいるかどうかの結果を見て、経験的に知るしかない。(=神のみ ぞ、知る)
* 略して、”人間万事、塞翁が馬”。( 注) ”人間”を”じんかん”と読まないで、あえてDNAを想定して”にんげん”と読む)